
Pada dasarnya, Fotosintesis merupakan proses penyusunan karbohidrat atau zat gula dengan menggunakan energi matahari. Matahari sebagai sumber energi utama bagi kehidupan di Bumi. Namun tidak semua organisma mampu secara langsung menggunakannya.
Hanya golongan tumbuhan dan beberapa jenis bakteri saja yang mampu menyerap energi matahari dan memanfaatkannya untuk fotosintesis. Melalui fotosintesis, tumbuhan menyusun zat makanan yaitu karbohidrat (pati/gula). Karena kemampuan menyusun makanannya sendiri inilah, tumbuhan disebut organisma ototrof. Bagaimana fotosintesis itu dipahami oleh para ahli biologi? Untuk itu, perlu kita ikuti percobaan – percobaan pada periode awal penemuannya.
Lihat Daftar Inti Pelajaran :
Pengertian dan Sejarah Fotosintesis
Fenomena fotosintesis telah digali sejak lama oleh para ilmuwan, khususnya bidang fisiologi tumbuhan. Joseph Priestley (1972), seorang ahli kimia Inggris menemukan bahwa tumbuhan mengeluarkan suatu gas yang membuat api lilin dapat menyala walaupun dalam tabung gelas yang tertutup.
Dalam sungkup tabung gelas tanpa tanaman, api lilin yang dinyalakan cepat padam. Namun setelah ke dalamnya disusupkan tanaman, pada beberapa hari kemudian ternyata lilin dapat dinyalakan lagi. Lilin tetap menyala selama “gas” dari tanaman itu masih ada. Pada waktu itu, Dia belum tahu bahwa gas itu adalah oksigen.
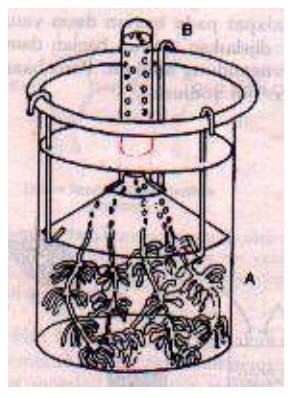
Dua ratus tahun kemudian, banyak peneliti tertarik untuk ikut menggali lebih lanjut dari temuan Priestley tersebut. Jan Ingenhousz (1779), ahli fisiologi dari German melakukan eksperimen dengan menggunakan tumbuhan air (Hydrila verticilata). Dari percobaannya ditunjukkan tiga hal penting, meliputi :
{kind=link}
Gambar 1. Percobaan Ingenhous
- Gas yang dikeluarkan oleh tumbuhan itu ternyata adalah O2,
- Cahaya matahari dibutuhkan untuk proses tersebut,
- Bagian yang berhijau daun saja yang mengeluarkan O2.
Seorang ahli botani dari Swiss, Jean Senebier menemukan bahwa CO2 juga dibutuhkan untuk fotosintesis. Peneliti lain, ahli kimia dan ahli fisiologi Swiss yaitu Nicholas de Saussure (1804) menunjukkan bahwa tanaman tumbuh dari air dan CO2 yang diserapnya. Sachs (1860) menunjukkan bahwa fotosintesis menghasilkan zat gula atau karbohidrat yang disebut amilum. Berdasar temuan-temuan itu maka pemahaman tentang fotosintesis menjadi semakin lengkap. Fotosintesis kemudian dirumuskan dalam persamaan reaksi kimia sbb :
Rumus Fotosintesis
n CO2 + n H2O + Energi Matahari —————–> [CH2O]n + nO2
klorofil zat gula
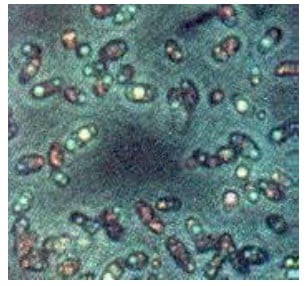
Semula orang mengira bahwa O2 yang dikeluarkan adalah berasal dari pemecahan gas CO2. Van Niel adalah orang pertama yang menyatakan bahwa O2 itu berasal dari pemecahan air. Hal itu didasarkan dari hasil temuannya tentang fotosintesis bakteri Sulfur.
{kind=link}
Gambar 2. Bakteri Sulfur
Dengan energi matahari, bakteri Sulfur ternyata juga mampu menyusun zat gula dari CO2 dan gas belerang (H2S), bukan dengan air (H2O) seperti pada tumbuhan. Bakteri ini melepaskan S, yang tentu berasal dari pemecahan H2S. Persamaan reaksinya dinyatakan sbb :
Energi matahari
CO2 + H2S —————–> [ CH2O ] + 2S + H2O
(zat gula)
Senada dengan hal itu, maka Van Niel menduga bahwa O2 yang dilepaskan pada fotosintesis tumbuhan adalah berasal dari pemecahan air (H2O). Tahun 1941, Ruben dan Kamen melakukan percobaan fotosintesis dengan menggunakan air bertanda. Pada air tersebut, komponen O-nya diberi tanda yang mudah dikenali dengan alat tertentu.
Dengan cara ini, Dia berhasil membuktikan bahwa “gas” yang dilepaskan itu adalah O2 yang bertanda. Oksigen itu tentu berasal dari pemecahan air bertanda. Pemecahan air dengan energi cahaya yang diserap oleh sel-sel daun yang berfotosintesis ini disebut fotolisis. Dengan demikian, persamaan fotosintesis yang lengkap adalah sbb :
n CO2 + 2n H2O + Energi Matahari —————–> [ CH2O ]n + n O2 + n H2O
klorofil ( zat gula )
Jadi, berdasarkan uraian di atas dapat kita tarik beberapa pengertian:
- Fotosintesis menggunakan energi matahari untuk menyusun zat gula sederhana.
- Zat gula disusun dari bahan dasar yaitu berupa H2O dan CO2.
- Fotosintesis menghasilkan bahan sisa berupa O2 dan H2
- Fotosintesis hanya dapat dilakukan oleh tumbuhan dan beberapa jenis bakteri. Fotosintesis menyusun zat gula dari air dan karbon dioksida (CO2), sehingga sering disebut pula asimilasi karbon.
Faktor yang Mempengaruhi Fotosintesis
Berikut faktor-faktor yang sangat berpengaruh terhadap fotosintesis, yaitu:
-
Respon fotosintesis terhadap intensitas cahaya
Cahaya mutlak dibutuhkan sebagai energi penggerak fotosintesis, namun demikian tingkat kebutuhan antar kelompok tumbuhan akan berbeda. Tidak pada setiap kondisi meningkatnya intensitas akan diikuti atau menyebabkan meningkatnya laju fotosintesis.
Cahaya matahari merupakan sumber energi utama fotosintesis. Albert Einstein menyebut energi matahari sebagai foton (kuantum). Cahaya mempengaruhi fotosintesis dalam tiga hal, yaitu intensitas cahaya, lama pencahayaan dan warna cahayanya.
Pada intensitas cahaya yang kurang, fotosintesisnya akan lambat. Sebaliknya, pada intensitas yang lebih tinggi, fotosintesis akan lebih cepat. Hal itu terjadi terutama pada tumbuh tumbuhan rumput, seperti jagung, tebu dan golongan rumput yang lain.
Tidak semua warna sinar dapat dimanfaatkan atau diserap secara optimal oleh tumbuhan. Klorofil menyerap semua warna sinar, kecuali sinar hijau. Sinar yang paling banyak diserap untuk fotosintesis adalah sinar merah (± 700 nm) dan biru (± 450nm). Jenis sinar yang lain juga diserap energinya walaupun dalam tingkat yang lebih rendah. Sinar hijau justru dipantulkan oleh klorofil, sehingga daun tampak berwarna hijau.
-
Umur jaringan daun
Selain faktor intensitas cahaya, umur daun juga sangat menentukan produktivitas daun dalam aktivitas fotosintesisnya. Kapasistas kemampuan daun melakukan fortosintesis berkembang seiring dengan perkembangan kedewasaan daun mencapai perkembangan dan pertumbuhan optimalnya.
Pada fase awal pertumbuhannya, daun muda masih menggantungkan asimilat dari daun dewasa lainnya (mengimport). Pada saat daun mencapai laju pertumbuhan optimum, produktivitasnya telah jauh meningkat, dan sebagian fotosintatnya telah mulai diekspor ke jaringan lain yang membutuhkan.
Kapasitas fotosintesis ini terus meningkat bersamaan dengan pencapaian kedewasaan organ daun. Terdapat hubungan interaktif antara perkembangan struktural daun (anatomi-morfologi) dan intensitas cahaya dengan perkembangan kapasitas fotosintetiknya. Tumbuhan yang tumbuh pada tempat dengan intensitas cahaya tinggi, daun berkembang dengan memadahi, sehingga kapasitas fotosintetiknya juga lebih besar.
-
Kadar CO2 dan O2
Konsentrasi CO2 sebagai salah satu prekursor atau bahan dasar asimilasi karbon tentu akan sangat berpengaruh pada produktivitas fotosintesisnya. Fotosintesis cenderung meningkat bila kadar CO2-nya lebih tinggi. Sebaliknya, keberadaan O2 justru akan menghambat fotosintesis. Tumbuhan menunjukkan kemampuannya dalam memfiksasi CO2 yang berbeda-beda.
Ada yang cepat dan ada juga yang lambat sesuai dengan tife tanaman tersebut. Oksigen merupakan salah satu produk samping dari fotosintesis. Namun demikian, kadar oksigen yang tinggi pada jaringan fotosintetik akan menghambat laju fotosintesis. Pada kondisi kadar oksigen yang semakin tinggi, laju fotosintesisnya secara signifikan menjadi semakin rendah.
Baca Juga :
Fotosintesis adalah | Pengertian dan Definisi
FOTOSINTESIS TUMBUHAN C3, C4, DAN CAM
Pengertian, Fungsi, Proses Fotosintesis
Reaksi Fotosintesis Gelap dan Terang
Tempat Terjadinya Fotosintesis
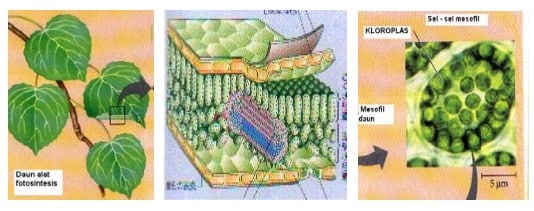
Kloroplas merupakan alat atau organela sel yang khas pada sel-sel daging daun.
{kind=link}
Gambar 3. Daun, Susunan Anatomi dan Kloroplas
Bentuk kloroplas bermacam-macam, tergantuing jenis tumbuhannya. Selain bulat atau lonjong, ada juga yang berbentuk pita. Sel sebagian besar tumbuhan tinggi umumnya mengandung antara 50 – 200 kloroplas. Kalau dilihat dari samping bentuknya seperti lensa dengan satu sisi/permukaan cembung dan permukaan lain cekung, datar atau cembung. Sumbu panjang kloroplas itu sering berukuran 5–10 μm. Dilihat dari atas kloroplas nampak sebagai elifotosistem.
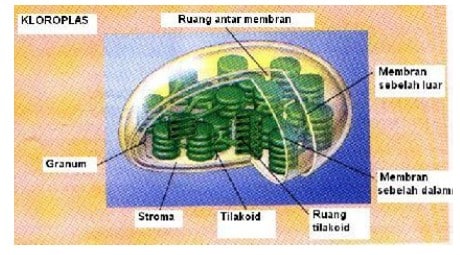{kind=link}
Gambar 4. Kloroplas dan Bagian-Bagiannya: Grana dan Stroma
Pada tumbuhan rendah dan terutama pada beberapa mikroorganisme, bentuknya sangat berbeda dari yang terlihat pada tumbuhan tinggi dan sering jumlahnya sedikit.
Sebagai contoh:
Euglena gracilis : kurang lebih 10 kloroplas/sel
Chlamydomonas : satu kloroplas/sel, berbentuk mangkuk
Spirogyra : satu kloroplas/sel, berbentuk pita yang memanjang di seluruh sel
Pada dasarnya, kloroplas dibatasi oleh dua sistem membran yaitu membran luar dan membran dalam, yang dipisahkan oleh ruang antar membran.
- Membran dalam
Membran dalam dihubungkan dengan suatu kompleks membran yaitu membran bagian dalam yang melintasi bagian dalam kloroplas. Dengan demikian, organel itu adalah suatu sistem tiga membran.
Bentuk membran bagian dalam yang paling umum adalah satu kantung yang dipipihkan yang disebut tilakoid. Tilakoid itu terdapat dalam stroma. Tumpukkan beberapa tilakoid disebut grana, sehingga masing-masing tilakoidnya disebut tilakoid grana. Tilakoid yang memanjang ke stroma disebut tilakoid stroma.
Bagian dalam tilakoid disebut lokulus. Membran-membran pada kloroplas membatasi tiga kompartemen yang terpisah yaitu ruang antar membran, stroma dan lokulus. Reaksi-reaksi fotosintesis bergantung cahaya berlangsung dalam tilakoid sedang reaksi asimilasi (fiksasi) CO2 terjadi dalam stroma.
Membran dalam bekerja sebagai pembatas fungsional antara sitosol dan stroma. Membran dalam tidak permeabel bagi sukrosa dan berbagai anion, misal di- dan trikarboksilat, fosfat dan senyawa-senyawa seperti nukleotida dan gula fosfat.
Membran dalam permeabel bagi CO2 dan asam-asam monokarboksilat tertentu, misal asam asetat, asam gliserat dan asam glikolat. Membran dalam kurang permeabel bagi asam amino. Membran dalam mengandung protein pembawa tertentu untuk mengangkut fosfat, fosfogliserat, dihidroksiaseton fosfat, dikarboksilat dan ATP.
Sistem membran bagian dalam yang terdapat dalam stroma membentuk suatu jalinan yang sangat kompleks. Membran tilakoid mengandung enzim lengkap untuk melaksanakan reaksi-reaksi fotosintesis yang bergantung cahaya. Membran tilakoid merupakan tempat klorofil, pembawa-pembawa elektron dan faktor-faktor yang menggabungkan transpor elektron dengan fosforilasi.
Stroma mengandung enzim-enzim yang penting untuk melaksanakan asimilasi CO2 dan mengubahnya menjadi karbohidrat. Beberapa macam partikel juga terdapat seperti butir pati, plastoglobulin yaitu tempat penyimpan lipida, plastokinon dan tokoforilkinon. Stroma juga mengandung ribosom dan DNA.
Membran tilakoid kira-kira 50% terdiri atas lipida, kurang lebih 10% dari padanya adalah fosfolipida. Lipida yang khas bagi klorofil adalah galaktolipida dan sulfolipida, yang masing-masing 45% dan 4% dari total lipida. Selain itu terdapat molekul-molekul lipida seperti klorofil, karotenoid dan plastokinon. Jumlah klorofil kira-kira 20% dari lipida total membran tilakoid.
- Membran luar
Membran luar kloroplas tumbuhan tinggi dipisahkan dari membran dalam oleh ruang kira-kira 10 nm. Membran tersebut permeabel bagi bermacam-macam senyawa dengan berat molekul rendah seperti nukleotida, fosfat organik, derivat-derivat fosfat, asam karboksilat dan sukrosa. Dengan demikian ruang antar membran mengandung molekul-molekul nutrien sitosol.
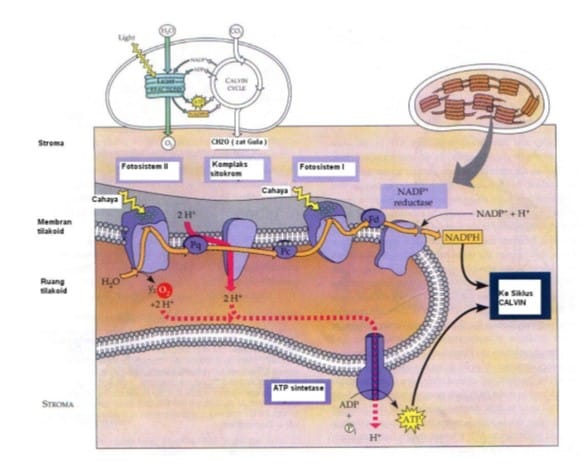
Proses Fotosintesis
Pada dasarnya, fotosintesis terjadi dalam dua tahapan. Kedua tahap itu berlangsung dalam kloroplas, namun pada dua bagian yang berbeda. Tahap I adalah proses penangkapan energi surya atau proses-proses yang bergantung langsung pada keberadaan cahaya.
Seluruh proses pada tahap ini disebut reaksi terang. Tahap II adalah proses-proses yang tidak bergantung langsung pada keberadaan cahaya. Proses-proses atau reaksi-reaksi pada tahap ini disebut reaksi gelap.
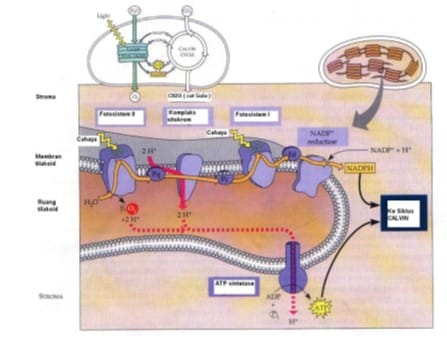{kind=link}
Gambar 5. Penangkapan energi surya (foton) dan aliran elektron dalam membran tilakoid grana (reaksi cahaya / Reaksi terang)
-
Reaksi Terang
Reaksi terang adalah proses untuk menghasilkan ATP dan reduksi NADPH2. Reaksi ini memerlukan molekul air dan cahaya matahari. Proses diawali dengan penangkapan foton oleh pigmen sebagai antena. Foton yang dimaksud adalah segmen spektrum yang paling penting bagi kehidupan yaitu pita sempit antara panjang gelombang sekitar 380 nm sampai 750 nm. Radiasi ini dikenal sebagai cahaya tampak, karena dapat dideteksi sebagai beraneka ragam warna oleh mata manusia.
Fotosistem adalah suatu unit yang mampu menangkap energi cahaya Matahari yang terdiri dari klorofil a, kompleks antena, dan akseptor elektron. Di dalam kloroplas terdapat beberapa macam klorofil dan pigmen lain, seperti klorofil a yang berwarna hijau muda, klorofil b berwarna hijau tua, dan karoten yang berwarna kuning sampai jingga. Pigmen-pigmen tersebut mengelompok dalam membran tilakoid dan membentuk perangkat pigmen yang berperan penting dalam fotosintesis.
Klorofil a berada dalam bagian pusat reaksi. Klorofil ini berperan dalam menyalurkan elektron yang berenergi tinggi ke akseptor utama elektron. Elektronini selanjutnya masuk ke sistemsiklus elektron. Elektron yang dilepaskan klorofil a mempunyai energi tinggi sebab memperoleh energi dari cahaya yang berasal dari molekul perangkat pigmen yang dikenal dengan kompleks antena.
Fotosistem sendiri dapat dibedakan menjadi dua, yaitu fotosistem I dan fotosistem II.
- Fotosistem I
Pada fotosistem I ini penyerapan energi cahaya dilakukan oleh klorofil a yang sensitif terhadap cahaya dengan panjang gelombang 700 nm sehingga klorofil a disebut juga P700. Energi yang diperoleh P700 ditransfer dari kompleks antena.
- Fotosistem II
Pada fotosistem II penyerapan energi cahaya dilakukan oleh klorofil a yang sensitif terhadap panjang gelombang 680 nm sehingga disebut P680. P680 yang teroksidasi merupakan agen pengoksidasi yang lebih kuat daripada P700. Dengan potensial redoks yang lebih besar, akan cukup elektron negatif untuk memperoleh elektron dari molekul-molekul air.
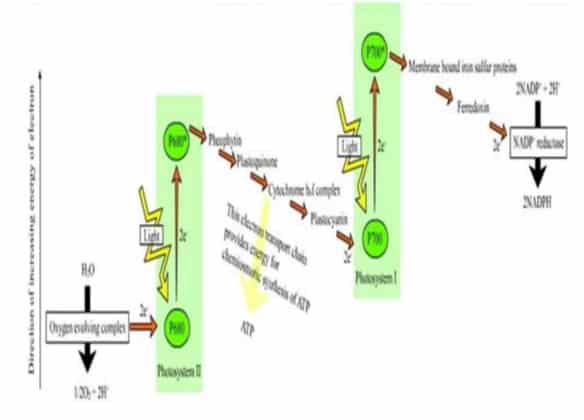{kind=link}
Gambar 6. Proses Fotosistem 1 dan 2
Mekanisme reaksi terang diawali dengan tahap dimana fotosistem II menyerap cahaya Matahari sehingga elektron klorofil pada fotosistem II tereksitasi dan menyebabkan muatan menjadi tidak stabil. Untuk menstabilkan kembali, fotosistem II akan mengambil elektron dari molekul H2O yang ada disekitarnya. Molekul air akan dipecahkan oleh ion mangan (Mn) yang bertindak sebagai enzim. Hal ini akan mengakibatkan pelepasan H+ di lumen tilakoid.
Dengan menggunakan elektron dari air, selanjutnya fotosistem II akan mereduksi plastokuinon (PQ) membentuk PQH2. Plastokuinon merupakan molekul kuinon yang terdapat pada membran lipid bilayer tilakoid. Plastokuinon ini akan mengirimkan elektron dari fotosistem II ke suatu pompa H+ yang disebut sitokrom b6-f kompleks.
Sitokrom b6-f kompleks berfungsi untuk membawa elektron dari fotosistem II ke fotosistem I dengan mengoksidasi PQH2 dan mereduksi protein kecil yang sangat mudah bergerak dan mengandung tembaga, yang dinamakan plastosianin (PC). Kejadian ini juga menyebabkan terjadinya pompa H+ dari stroma ke membran tilakoid.
Elektron dari sitokrom b6-f kompleks akan diterima oleh fotosistem I. Fotosistem ini menyerap energi cahaya terpisah dari fotosistem II, tapi mengandung kompleks inti terpisahkan, yang menerima elektron yang berasal dari H2O melalui kompleks inti Fotosistem II lebih dahulu. Sebagai sistem yang bergantung pada cahaya, Fotosistem I berfungsi mengoksidasi plastosianin tereduksi dan memindahkan elektron ke protein Fe-S larut yang disebut feredoksin.
Selanjutnya elektron dari feredoksin digunakan dalam tahap akhir pengangkutan elektron untuk mereduksi NADP dan membentuk NADPH. Reaksi ini dikatalisis dalam stroma oleh enzim feredoksin-NADP+ reduktase.
Ion H+ yang telah dipompa ke dalam membran tilakoid akan masuk ke dalam ATP sintase. ATP sintase akan menggandengkan pembentukan ATP dengan pengangkutan elektron dan H+ melintasi membran tilakoid. Masuknya H+ pada ATP sintase akan membuat ATP sintase bekerja mengubah ADP dan fosfat anorganik (Pi) menjadi ATP.
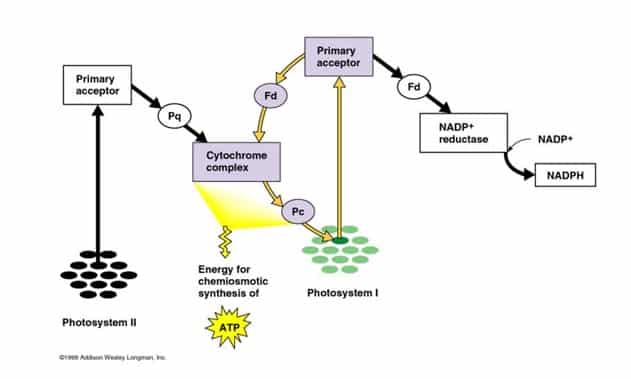
Reaksi Siklik dan NonSiklik
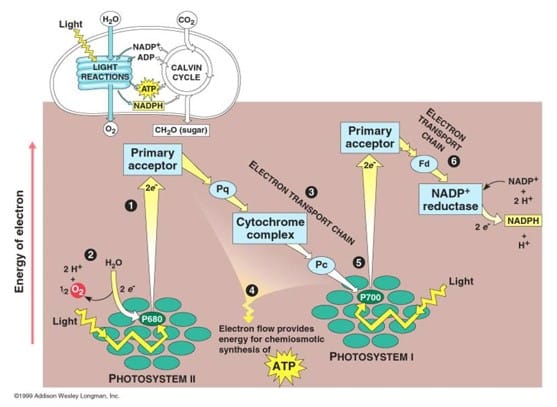
Reaksi terang memiliki dua bentuk: siklus dan nonsiklus. Pada reaksi nonsiklus, foton diserap pada kompleks antena fotosistem II penyerap cahaya oleh klorofil dan pigmen aksesoris lainnya. Ketika molekul klorofil pada inti pusat reaksi fotosistem II memperoleh energi eksitasi yang cukup dari pigmen antena yang berdekatan dengannya, satu elektron akan dipindahkan ke molekul penerima elektron, yaitu feopftin, melalui sebuah proses yang disebut pemisahan tenagaterfotoinduksi. Elektron ini dipindahkan melalui rangkaian transport elektron, yang disebut skema Z, yang pada awalnya berfungsi untuk menghasilkan potensi kemiosmosis di sepanjang membran.
Satu enzim sintase ATP menggunakan potensi kemisomosis untuk menghasilkan ATP selama fotofosforilasi, sedangkanNADPH adalah produk dari reaksi redoksterminal pada skema Z. Elektron masuk ke molekul klorofil pada fofosistem II. Elektron ini tereksitasi karena cahaya yang diserap oleh fotosistem. Pembawa elektron kedua menerima elektron, yang lagi-lagi dilewatkan untuk menurunkan energi penerim elektron. Energi yang dihasilkan oleh penerima elektron digunakan untuk menggerakan ion hidrogen di sepanjang membran tilakoid sampai ke dalam lumen. Elektron digunakan untuk mereduksi koenzim NADP, yang memiliki fungsi pada reaksi terang.
Reaksi siklus mirip dengan nonsiklus, namun berbeda pada bentuknya karena hanya menghasilkan ATP, dan tidak ada NADP (NADPH) tereduksi yang dihasilkan. Reaksi siklus hanya berlangsung pada fotosistem I. Setelah elektron dipindahkan dari fotosistem, elektron digerakkan melewati molekul penerima elektron dan dikembalikan ke fotosistem I, yang dari sanalah awalnya elektron dikeluarkan, sehingga reaksi ini diberi nama reaksi siklus.
{kind=link}
Gambar 7. Reaksi Non Siklik
{kind=link}
Gambar 8. Reaksi Siklik
-
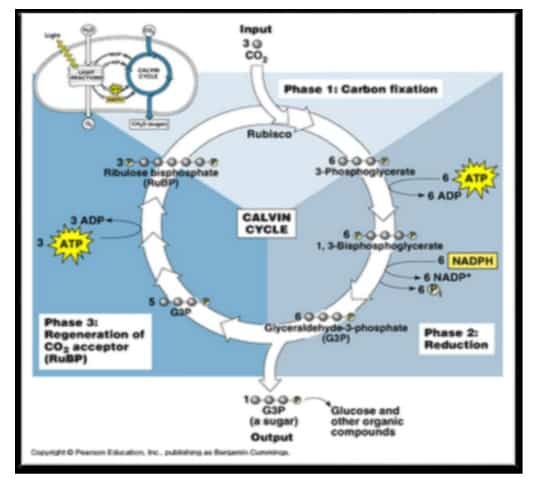
Reaksi Gelap (Siklus Calvin)
{kind=link}
Gambar 9. Siklus Calvin
Pada proses ini terjadi pengikatan karbondioksida di dalam daun. Siklus ini menggunakan ATP dan NADPH sebagai sumber energi dan NADPH sebagai tenaga pereduksi pembuatan gula. Karbohidrat yang dihasilkan langsung dari siklus Calvin sebenarnya bukan glukosa melainkan gula berkarbon tiga yang disebut gliseraldehida 3 fosfat (G3P). Berdasarkan Campbell (2002) daur Calvin dapat dibagi ke dalam 3 fase sebagai berikut:
- Pengikatan (fiksasi) CO2
CO2 diikat oleh senyawa ribulosa bifosfat (RuBP) untuk membentuk senyawa C-6 yang akan terurai menjadi dua molekul 1,3 bifosfogliserat. Enzim yang berperan dalam fiksasi CO2 adalah RuBP karboksilase atau rubisko.
- Reduksi
Molekul 1,3 bifosfogliserat akan diubah menjadi G3P dengan menambahkan 2 elektron dari 2 NADPH. Siklus ini harus berjalan 3 kali sehingga terbentuk 6 molekul G3P.
- Pembentukan RuBP (Ribolusa Bifosfat)
Pada tahap ini pembentukan RuBP dari 5 molekul G3P yang membutuhkan 3 ATP. Jadi untuk membuat 1 G3P dibutuhkan 9 ATP dan 6 NADPH. G3P dapat diubah menjadi dihidroksiaseton fosfat. Untuk membentuk 1 molekul glukosa dibutuhkan siklus Calvin yang berdaur ulang selama 6 kali, dan ditangkap 6 molekul.
Tipe Tipe Fotosintesis
Berdasarkan tipe fotosintesis, tumbuhan dibagi ke dalam tiga kelompok besar, yaitu C3, C4, dan CAM (crassulacean acid metabolism). Tumbuhan C4 dan CAM lebih adaptif di daerah panas dan kering dibandingkan dengan tumbuhan C3. Namun tanaman C3 lebih adaptif pada kondisi kandungan CO2 atmosfer tinggi. Sebagian besar tanaman pertanian, seperti gandum, kentang, kedelai, kacang-kacangan, dan kapas merupakan tanaman dari kelompok C3. Tanaman C3 dan C4 dibedakan oleh cara mereka mengikat CO2 dari atmosfir dan produk awal yang dihasilkan dari proses assimilasi.
- Tipe tanaman C3
Pada tanaman C3, enzim yang menyatukan CO2 dengan RuBP (RuBP merupakan substrat untuk pembentukan karbohidrat dalam proses fotosintesis) dalam proses awal assimilasi, juga dapat mengikat O2 pada saat yang bersamaan untuk proses fotorespirasi ( fotorespirasi adalah respirasi,proses pembongkaran karbohidrat untuk menghasilkan energi dan hasil samping, yang terjadi pada siang hari) . Jika konsentrasi CO2 di atmosfir ditingkatkan, hasil dari kompetisi antara CO2 dan O2 akan lebih menguntungkan CO2, sehingga fotorespirasi terhambat dan assimilasi akan bertambah besar.
- Tipe tanaman C4
Pada tanaman C4, CO2 diikat oleh PEP (enzym pengikat CO2 pada tanaman C4) yang tidak dapat mengikat O2 sehingga tidak terjadi kompetisi antara CO2 dan O2. Lokasi terjadinya assosiasi awal ini adalah di sel-sel mesofil (sekelompok sel-sel yang mempunyai klorofil yang terletak di bawah sel-sel epidermis daun). CO2 yang sudah terikat oleh PEP kemudian ditransfer ke sel-sel “bundle sheath” (sekelompok sel-sel di sekitar xylem dan phloem) dimana kemudian pengikatan dengan RuBP terjadi.
Karena tingginya konsentasi CO2 pada sel-sel bundle sheath ini, maka O2 tidak mendapat kesempatan untuk bereaksi dengan RuBP, sehingga fotorespirasi sangat kecil and G sangat rendah, PEP mempunyai daya ikat yang tinggi terhadap CO2, sehingga reaksi fotosintesis terhadap CO2 di bawah 100 m mol m-2 s-1 sangat tinggi.
Laju assimilasi tanaman C4 hanya bertambah sedikit dengan meningkatnya CO2. Sehingga, dengan meningkatnya CO2 di atmosfir, tanaman C3 akan lebih beruntung dari tanaman C4 dalam hal pemanfaatan CO2 yang berlebihan. Contoh tanaman C3 antara lain : kedele, kacang tanah, kentang, sedangkan contoh tanaman C4 adalah : jagung, sorgum dan tebu.
Sintesis Tipe Tanaman C3, C4, dan CAM, yaitu:
- Sintesis C3
Sintesis C3 diawali dengan fiksasi CO2, yaitu menggabungkan CO2 dengan sebuah molekul akseptor karbon. Akan tetapi didalam sintesis C3, CO2 difiksasi ke gula berkarbon 5, yaitu ribulosa bifosfat (RuBP) oleh enzim karboksilase RuBP (rubisko). Molekul berkarbon 6 yang berbentuk tidak stabil dan segera terpisah menjadi 2 molekul fosfogliserat (PGA). Molekul PGA merupakan karbohidrat stabil berkarbon 3 yang pertama kali terbentuk sehingga cara tersebut dinamakan sintesis C3.
Molekul PGA bukan molekul berenergi tinggi. Dua molekul PGA mengandung energy yang lebih kecil dibandingkan dengan satu molekul RuBP. Hal tersebut menjelaskan alasan fiksasi CO2 berlangsung secara spontan dan tidak memerlukan energy dari reaksi cahaya. Untuk mensintesis molekul berenergi tinggi, energy dan electron dari ATP maupun NADPH hasil reaksi terang digunakan untuk mereduksi tiap PGA menjadi fosfogliseraldehida (PGAL).
Dua molekul PGAL dapat membentuk satu glukosa. Siklus Calvin telah lengkap bila pembentukan glukosa disertai dengan generasi RuBP. Satu molekul CO2 yang tercampur menjadi enam molekul CO2. Ketika enam molekul CO2 bergabung dengan enam molekul RuBP dihasilkan satu glukosa dan enam RuBP sehingga siklus dapat dimulai lagi. Contoh tanaman: legum (polong-polongan), gandum, padi.
- Sintesis C4
Pada jenis tumbuhan yang hidup di daerah panas seperti jagung, tebu, rumput-rumputan, memiliki kebiasaan saat siang hari mereka tidak membuka stomatanya secara penuh untuk mengurangi kehilangan air melalui evaporasi/transpirasi. Ini berakibat terjadinya penurunan jumlah CO2 yang masuk ke stomata. Logikanya hal ini menghambat laju fotosintesis. Ternyata para tumbuhan ini telah mengembangkan cara yang cerdas untuk menjaga agar laju fotosintesis tetap normal meskipun stomata tidak membuka penuh.
Perbedaan tanaman C3 dan C4 adalah ada pada mekanisme fiksasi CO2. Pada tumbuhan C-4 karbondioksida pertamakali akan diikat oleh senyawa yang disebut PEP (phosphoenolphyruvate / fosfoenolpiruvat) dengan bantuan enzim PEP karboksilase dan membentuk oksaloasetat, suatu senyawa 4-C. Itu sebabnya kelompok tumbuhan ini disebut tumbuhan C-4 atau C-4 pathway. PEP dibentuk dari piruvat dengan bantuan enzim piruvat fosfat dikinase. Berbeda dengan rubisco, PEP sangat lemah berikatan dengan O2.
Ini berarti bisa menekan terjadinya fotorespirasi sekaligus mampu menangkap lebih banyak CO2 sehingga bisa meningkatkan laju produksi glukosa. Pengikatan CO2 oleh PEP tersebut berlangsung di sel-sel mesofil (daging daun). Oksaloasetat yang terbentuk kemudian akan direduksi karena menerima H+ dari NADH dan berubah menjadi malat, kemudian ditransfer menuju ke sel seludang pembuluh (bundle sheath cells) melalui plasmodesmata. Sel-sel seludang pembuluh adalah kelompok sel yang mengelilingi jaringan pengangkut xilem dan floem.
Di dalam sel-sel seludang pembuluh malat akan dipecah kembali menjadi CO2 yang langsung memasuki siklus Calvin-Benson, dan piruvat dikembalikan lagi ke sel-sel mesofil. Hasil dari siklus Calvin-Benson adalah molekul glukosa yang kemudian ditranspor melalui pembuluh floem.
Dari uraian di atas kita tahu bahwa fiksasi CO2 pada tumbuhan C-4 berlangsung dalam dua langkah.
- CO2 diikat oleh PEP menjadi oksaloasetat dan berlangsung di sel-sel mesofil.
- CO2 diikat oleh rubisco menjadi APG di sel seludang pembuluh.
Ini menyebabkan energi yang digunakan untuk fiksasi CO2 lebih besar, memerlukan 30 molekul ATP untuk pembentukan satu molekul glukosa. Sedangkan pada tumbuhan C-3 hanya memerlukan 18 molekul ATP. Namun demikian besarnya kebutuhan ATP untuk fiksasi CO2 pada tumbuhan C-4 sebanding dengan besarnya hasil produksi glukosa karena dengan cara tersebut mampu menekan terjadinya fotorespirasi yang menyebabkan pengurangan pembentukan glukosa. Itu sebabnya kelompok tumbuhan C-4 dikenal efektif dalam fotosintesis.
- Sintesis CAM
Tumbuhan lain yang tergolong sukulen (penyimpan air) misalnya kaktus dan nanas memiliki adaptasi fotosintesis yang berbeda lagi. Tidak seperti tumbuhan umumnya, kelompok tumbuhan ini membuka stomata pada malam hari dan menutup pada siang hari. Stomata yang menutup pada siang hari membuat tumbuhan mampu menekan penguapan sehingga menghemat air, tetapi mencegah masuknya CO2.
Saat stomata terbuka pada malam hari, CO2 di sitoplasma sel-sel mesofil akan diikat oleh PEP dengan bantuan enzim PEP karboksilase sehingga terbentuk oksaloasetat kemudian diubah menjadi malat (persis seperti tumbuhan C-4). Selanjutnya malat yang terbentuk disimpan dalam vakuola sel mesofil hingga pagi hari. Pada siang hari saat reaksi terang menyediakan ATP dan NADPH untuk siklus Calvin-Benson, malat dipecah lagi menjadi CO2 dan piruvat. CO2 masuk ke siklus Calvin-Benson di stroma kloroplas, sedangkan piruvat akan digunakan untuk membentuk kembali PEP.
Manfaat Hasil Fotosintesis
Manfaat Fotosintesis Bagi Tumbuhan dan Kehidupan
-
Bagi Tumbuhan
Zat gula hasil fotosintesis akan digunakan untuk berbagai kepentingan tubuh tumbuhan. Sebagian zat gula akan dirombak untuk menghasilkan energi. Energi sangat dibutuhkan untuk berbagai aktivitas tubuh. Sebagian akan digunakan untuk membangun atau membentuk tubuh tumbuhan.
Tumbuhan butuh tumbuh, berkembang, membentuk anakan atau bertunas, membentuk bunga, buah, biji, dsb. Sebagian akan dijadikan bahan baku untuk menyusun zat-zat penting lain yang dibutuhkan. Misalnya, protein, lemak dan vitamin. Sebagian yang lain akan ditimbun dalam jaringan penimbunan, misalnya dalam bentuk ubi, umbi, buah dan biji.
- Bagi kehidupan
Salah satu manfaat fotosintesis selain untuk tumbuhan melakukan metabolisme adalah yaitu fotosintesis mengilhami para peneliti untuk membuat sel surya yang menghadapi sistem fotosintesis tersebut.
Sel surya atau juga sering disebut fotovoltaik adalah divais yang mampu mengkonversi langsung cahaya matahari menjadi listrik. Sel surya bisa disebut sebagai pemeran utama untuk memaksimalkan potensi sangat besar energi cahaya matahari yang sampai kebumi, walaupun selain dipergunakan untuk menghasilkan listrik, energi dari matahari juga bisa dimaksimalkan energi panasnya melalui sistem solar thermal.
Cara Kerja Sel Surya
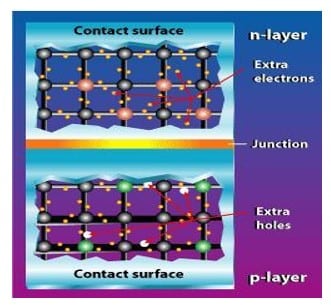
Sel surya konvensional bekerja menggunakan prinsip p-n junction, yaitu junction antara semikonduktor tipe-p dan tipe-n. Semikonduktor ini terdiri dari ikatan-ikatan atom yang dimana terdapat elektron sebagai penyusun dasar. Semikonduktor tipe-n mempunyai kelebihan elektron (muatan negatif) sedangkan semikonduktor tipe-p mempunyai kelebihan hole (muatan positif) dalam struktur atomnya.
Kondisi kelebihan elektron dan hole tersebut bisa terjadi dengan mendoping material dengan atom dopant. Sebagai contoh untuk mendapatkan material silikon tipe-p, silikon didoping oleh atom boron, sedangkan untuk mendapatkan material silikon tipe-n, silikon didoping oleh atom fosfor. Ilustrasi dibawah menggambarkan junction semikonduktor tipe-p dan tipe-n.
{kind=link}
Gambar 10. Junction antara semikonduktor tipe-p (kelebihan hole) dan tipe-n (kelebihan elektron).
Peran dari p-n junction ini adalah untuk membentuk medan listrik sehingga elektron (dan hole) bisa diekstrak oleh material kontak untuk menghasilkan listrik. Ketika semikonduktor tipe-p dan tipe-n terkontak, maka kelebihan elektron akan bergerak dari semikonduktor tipe-n ke tipe-p sehingga membentuk kutub positif pada semikonduktor tipe-n, dan sebaliknya kutub negatif pada semikonduktor tipe-p.
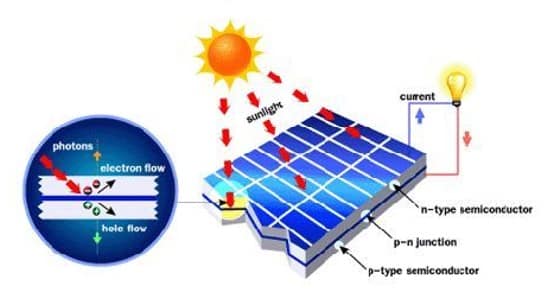
Akibat dari aliran elektron dan hole ini maka terbentuk medan listrik yang mana ketika cahaya matahari mengenai susuna p-n junction ini maka akan mendorong elektron bergerak dari semikonduktor menuju kontak negatif, yang selanjutnya dimanfaatkan sebagai listrik, dan sebaliknya hole bergerak menuju kontak positif menunggu elektron datang, seperti diilustrasikan pada gambar dibawah.
{kind=link}
Gambar 11. Ilustrasi cara kerja sel surya dengan prinsip p-n junction.